Ir al contenido principal
Los mecanismos de la evolución

Hay dos mecanismos básicos de cambio evolutivo: la selección natural y la deriva genética. La selección natural favorece a los genes que mejoran la capacidad de supervivencia y reproducción del organismo. La deriva genética es el cambio aleatorio en la frecuencia de los alelos, provocado por muestreo aleatorio de los genes de una generación a la siguiente. El flujo genético es la transferencia de genes dentro de una población o entre poblaciones. La importancia relativa de la selección natural y de la deriva genética en una población varía dependiendo de la fuerza de la selección y del tamaño poblacional efectivo, que es el número de ejemplares de esa población capaces de reproducirse.154 La selección natural suele predominar en las poblaciones grandes, mientras que la deriva genética predomina en las pequeñas. El predominio de la deriva genética en poblaciones pequeñas puede llevar incluso a la fijación de mutaciones ligeramente deletéreas.155 Como resultado de ello, los cambios en el tamaño de una población pueden influir significativamente en el curso de la evolución. Los cuellos de botella en las poblaciones —situaciones que llevan a un drástico descenso temporario del tamaño efectivo— determinan una pérdida o erosión de la variabilidad genética y conllevan, por lo tanto, a la formación de poblaciones genéticamente más uniformes. Los cuellos de botella también pueden ser el resultado de alteraciones en el flujo genético, como una migración reducida, la expansión a nuevos hábitats, o una subdivisión de la población.
Selección natural
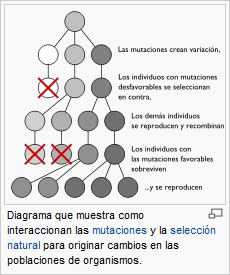
La selección natural es el proceso por el cual las mutaciones genéticas que mejoran la capacidad reproductiva se vuelven, y permanecen, cada vez más frecuentes en las sucesivas generaciones de una población. Se la califica a menudo de «mecanismo autoevidente», pues es la consecuencia necesaria de tres hechos simples: (a) dentro de las poblaciones de organismos hay variación heredable, (b) los organismos producen más descendientes de los que pueden sobrevivir, y (c) tales descendientes tienen diferentes capacidades para sobrevivir y reproducirse.
El concepto central de la selección natural es la aptitud biológica de un organismo.157 La aptitud, ajuste o adecuación se trata de la medida de la contribución genética de un organismo a la generación siguiente.157 Sin embargo, la aptitud no es simplemente igual al número total de descendientes de un determinado organismo, ya que también cuantifica la proporción de generaciones posteriores que llevan los genes de ese organismo.158 Por ejemplo, si un organismo puede sobrevivir y reproducirse pero sus descendientes son demasiado pequeños o enfermizos como para llegar a la edad reproductiva, la contribución genética de ese organismo a las futuras generaciones será muy baja y, por ende, su aptitud también lo es.

Por consiguiente, si un alelo aumenta la aptitud más que otros alelos del mismo gen, con cada generación el alelo será más común dentro de la población. Se dice que tales rasgos son «seleccionados a favor». Ejemplos de rasgos que pueden aumentar la aptitud son una mejora de la supervivencia o una mayor fecundidad. En cambio, la menor aptitud causada por un alelo menos beneficioso o deletéreo hace que el alelo sea cada vez más raro y se dice que es «seleccionado en contra».159 Hay que subrayar que la aptitud de un alelo no es una característica fija, si el ambiente cambia, los rasgos que antes eran neutros o nocivos pueden ser beneficiosos, y viceversa.160 Por ejemplo, la polilla Biston betularia presenta dos colores, uno claro denominado forma typica y otro oscuro llamado forma carbonaria. La forma typica, como su nombre indica, es la más frecuente en esta especie. No obstante, durante la revolución industrial en el Reino Unido los troncos de muchos de los árboles en los que las polillas se posaban resultaron ennegrecidos por el hollín, lo que les proporcionaba una ventaja a las polillas de color oscuro para pasar desapercibidas de los depredadores. Esto dio a las polillas de la forma melanica una mayor oportunidad de sobrevivir para producir más descendientes de color oscuro. En sólo cincuenta años luego de que la primer polilla melánica fuese descubierta, casi la totalidad de las polillas del área industrial de Manchester eran oscuras. Este proceso fue revertido por el efecto de la «Ley de Aire Limpio» (Clean Air Act) de 1956 por la cual se redujo la polución industrial y las polillas oscuras, más fácilmente visibles por los depredadores, volvieron a ser escasas nuevamente.
Sin embargo, aunque la dirección de la selección cambie, los rasgos que se hubiesen perdido en el pasado pueden no reobtenerse nuevamente de forma idéntica —situación que describe la Ley de Dollo o «Ley de la irreversibilidad evolutiva»—.162 De acuerdo con esta hipótesis, una estructura u órgano que se ha perdido o descartado en él durante el transcurso de la evolución no volverá a aparecer en ese mismo linaje de organismos.163 164 Según Richard Dawkins, esta hipótesis es «una declaración sobre la improbabilidad estadística de seguir exactamente la misma trayectoria evolutiva dos veces o, de hecho, una misma trayectoria particular en ambas direcciones».
Dentro de una población, la selección natural para un determinado rasgo que varía en forma continua, como la altura, se puede categorizar en tres tipos diferentes. El primero es la «selección direccional», que es un cambio en el valor medio de un rasgo a lo largo del tiempo; por ejemplo, cuando los organismos cada vez son más altos.166 En segundo lugar se halla la «selección disruptiva» que es la selección de los valores extremos de un determinado rasgo, lo que a menudo determina que los valores extremos sean más comunes y que la selección actúe en contra del valor medio. Esto implicaría que los organismos bajos y altos tengan una ventaja, pero los de altura media no. Finalmente, en la «selección estabilizadora», la selección actúa en contra de los valores extremos, lo que determina una disminución de la varianza alrededor del promedio y una menor variabilidad de la población para ese carácter en particular.156 167 Esto haría, por ejemplo, que todos los organismos de una población, paulatinamente, adquirieran una altura similar.
Un tipo especial de selección natural es la selección sexual, que es la selección a favor de cualquier rasgo que aumente el éxito reproductivo haciendo aumentar el atractivo de un organismo ante parejas potenciales.168 Los rasgos que evolucionaron mediante la selección sexual son especialmente prominentes en los machos de algunas especies, aunque ciertos rasgos —tales como cuernos voluminosos, cantos de apareamiento o colores brillantes— puedan atraer a los predadores, reduciendo las posibilidades de supervivencia de los machos.169 No obstante, esta desventaja reproductiva se compensa por un mayor éxito reproductivo de los machos que presentan estos rasgos sexualmente seleccionados.170
Un área de estudio activo es la denominada «unidad de selección»; se ha dicho que la selección natural actúa a nivel de genes, células, organismos individuales, grupos de organismos e incluso especies.171 172 Ninguno de estos modelos es mutuamente exclusivo, y la selección puede actuar en múltiples niveles a la vez.173 Por ejemplo, debajo del nivel del individuo, hay genes denominados transposones que intentan replicarse en todo el genoma.174 La selección por sobre el nivel del individuo, como la selección de grupo, puede permitir la evolución de la cooperación.
Deriva genética

La deriva genética es el cambio en la frecuencia de los alelos entre una generación y la siguiente, y tiene lugar porque los alelos de la descendencia son una muestra aleatoria de los padres, y por el papel que juega el azar en la hora de determinar si un ejemplar determinado sobrevivirá y se reproducirá.95 En términos matemáticos, los alelos están sujetos a errores de muestreo. Como resultado de ello, cuando las fuerzas selectivas están ausentes o son relativamente débiles, la frecuencia de los alelos tiende a «derivar» hacia arriba o hacia abajo aleatoriamente (en un paseo aleatorio). Esta deriva se detiene cuando un alelo se convierte finalmente fijado, es decir, o bien desaparece de la población, o bien sustituye totalmente el resto de genes. Así pues, la deriva genética puede eliminar algunos alelos de una población simplemente debido al azar. Incluso en la ausencia de fuerzas selectivas, la deriva genética puede hacer que dos poblaciones separadas que empiezan con la misma estructura genética se separen en dos poblaciones divergentes con un conjunto de alelos diferentes.
El tiempo necesario para que un alelo quede fijado por la deriva genética depende del tamaño de la población; la fijación tiene lugar más rápido en poblaciones más pequeñas.177 La medida precisa de las poblaciones que es importante en este caso recibe el nombre de tamaño poblacional efectivo, que fue definida por Sewall Wright como el número teórico de ejemplares reproductivos que presenten el mismo grado observado de consanguinidad.
Aunque la selección natural es responsable de la adaptación, la importancia relativa de las dos fuerzas, selección natural y deriva genética, como impulsoras del cambio evolutivo en general es actualmente un campo de investigación en la biología evolutiva.178 Estas investigaciones fueron inspiradas por la teoría neutralista de la evolución molecular, que postula que la mayoría de cambios evolutivos son el resultado de la fijación de mutaciones neutras, que no tienen ningún efecto inmediato sobre la aptitud de un organismo.179 Por tanto, en este modelo, la mayoría de los cambios genéticos en una población son el resultado de una presión de mutación constante y de deriva genética.



Comentarios
Publicar un comentario